An easy example is the body shapes of sharks (fish), cetaceans (mammals: whales, dolphins, etc), and ichthyosaurs (porpoise-like reptiles): all evolved fairly similar adaptions for swimming, although a notable difference is the tailfins of cetaceans, giving rise to an up-and-down tail movement, rather than side-to-side for the others. A ready observation would be that they show similar vertebrate adaptions to the similar demands of the environment.
There are so many other examples: similar-shaped and/or similar-functioned animals around the world. Try unrelated fish in the arctic and antarctic waters each with an equivalent anti-freeze-like chemical in their blood, with genetic origins quite unconnected.
Try re-invention of the eye several times. Albeit the squid's, for example, is better engineered than ours, since our retinal nerve endings emerge between lens and retina, making for less efficient vision that the squid's whose nerve endings sensibly emerge behind the retina. Moving ahead of myself in Richard Dawkins' The Ancestor's Tale (currently at p344), Dawkins mentions on p602 an expert in comparative zoology of eyes, Professor Michael Land, who identifies nine independent principles of optical mechanics, "each of which has evolved more than once".
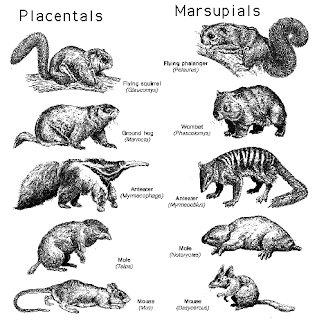
Try the proliferation of parallels between eutherian (placental) mammals and metatherian (marsupial) equivalents. Thylacine and timber wolf skulls, for example are said to be nearly identical, despite being completely unrelated.
It sounds like a rash of grossly unlikely coincidences, ready-made for creationists to fan argumentative flames. Yet in some ways it makes perfect sense: evolution is about species expanding to fill environmental niches, and
a) Over many times and locations, there are sets of all but identical niches
b) What developed and survived was what was most successful in the environment, so the directive forces of nature operated in a similar way on what was often a relatively similar genetic material.
This is obviously quite a source of frustration for taxonomy. Whereas in the relatively recent past, classification was based largely on morphology (body shape and features), modern molecular analysis has demonstrated that many of the connections drawn in the past were examples of convergence.
Dawkins also describes three different moles. The African golden mole (Chrysochloridae) was grouped with the Eurasian mole (Talpidae) in the defunct order Insectivora on the basis of such close similarity as burrowing machines: forepaws modified as spades; atrophied eyes (superfluous underground) and no visible ears. Then there's the marsupial mole: Wikipedia says they are so similar to golden moles that they were once thought to be related despite the marsupial/placental divide. Of course, all three are now classified quite separately; molecular analysis has in fact torn down the whole disputed Insectivora order altogether.
Interesting to note convergent evolution depicted as a criticism of Stephen Jay Gould. One of Gould's consistent themes was the contingent nature of evolution: how randomness played such a role that any slight re-alignment would have resulted in totally different outcomes.
I don't see these concepts as being entirely in opposition. Gould's theme plays out on a much larger tapestry than the micro-evolutionary outcomes. Convergence doesn't necessarily mean, for example, that had the K-T meteor not wiped out non-avian dinosaurs, reptilian humans would have evolved. In the absence of that meteor, dinosaurs could have remained successful in their niches for many millions more years given no other environmental pressures.
I suspect that certain paths are available only for species at a certain level of complexity; yet evolution is not specifically directional towards complexity (merely towards variation in complexity)...
How convergence does and doesn't work, what it does do and what it doesn't, is a fascinating area of study.
Reference
Dawkins, R (2004): The Ancestor's Tale. Phoenix, London.













